![Siklus Hidup Dan Habitat Cryptosporidium SP [PDF]](https://pdfs.asia/img/200x200/siklus-hidup-dan-habitat-cryptosporidium-sp.jpg)
4 0 199 KB
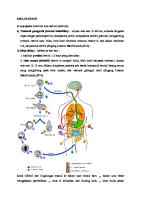
Siklus Hidup dan Habitat Cryptosporidium sp. Pada Manusia Cryptosporidium sp. pada manusia tidak hanya di usus halus, tetapi juga di organ-organ lain seperti faring, esofagus, lambung, duodenum, yeyunum, ileum, appendiks, kolon, rektum, kandung empedu dan saluran pankreas. Infeksi terberat ditemukan di 3 yeyunum. Pada individu immunokompeten, parasit biasanya terbatas di usus halus sedangkan pada individu imunokompromis dapat ditemukan sepanjang saluran pencernaan, selain itu juga dapat ditemukan pada sistem hepatobiliary dan saluran pernafasan. Cryptosporidium sp. di dalam usus, melekatkan diri di permukaan sel epitel pada “brush border”. Perkembangan parasit terjadi di dalam vakuola parasitoforus. Parasit ini intraseluler, tetapi ekstrasitoplasmik pada brush border sel hospes. Pada pangkal perlekatan antara parasit dan mikrovili ada daerah yang mengalami fusi. Daerah tersebut mempunyai struktur sangat tipis dan berbentuk lipatan-lipatan yang disebut feder organella yaitu tempat terjadinya aliran nutrisi dari sel hospes ke parasit Cryptosporidium sp. Infeksi Cryptosporidium sp. terjadi bila tertelan ookista matang yang dikeluarkan oleh tinja hospes terinfeksi. Masa prepaten yaitu waktu antara infeksi dan pengeluaran ookista selama 4 – 22 hari untuk manusia dan dapat lebih dari 30 hari pada individu imunokompeten. Siklus hidup Cryptosporidium sp. terdiri dari enam tahap perkembangan. Siklus seksual dan aseksual berlangsung dalam satu tubuh hospes (monoxenous) di saluran pencernaan. Siklus hidupnya dimulai dengan ekskistasi yang terjadi di traktus gastrointestinal atas. Sporozoit keluar dari ookista dan menempel pada sel epitel usus yang kemudian menjadi trofozoit. Trofozoit dan stadium-stadium selanjutnya hanya terdapat pada permukaan sel epitel, tidak masuk ke dalam sitoplasma epitel tersebut. Trofozoit mengalami skizogoni dengan tiga kali pembelahan inti dan terbentuk skizon generasi pertama yang mengandung delapan merozoit. Skizon pecah dan keluar delapan merozoit yang dapat menginfeksi sel epitel lain. Merozoit generasi pertama mengalami skizogoni lagi dengan dua kali pembelahan inti sehingga terbentuk skizon generasi kedua yang mengandung empat merozoit. Skizon tersebut pecah dan keluar merozoit yang dapat menginfeksi sel epitel lain, selain itu merozoit tersebut
ada yang tumbuh menjadi makrogametosit dan mikrogametosit. Makrogametosit mengalami
perkembangan
dan
berubah
menjadi
makrogamet.
Satu
makrogametosit akan berubah menjadi satu makrogamet. Mikrogametosit mengalami pembelahan inti beberapa kali dan berubah menjadi beberapa mikrogamet (12-16 mikrogamet). Satu mikrogamet akan membuahi satu makrogamet sehingga terbentuk zigot. Zigot akan berkembang menjadi ookista yang mengandung empat sporozoit. Siklus hidup Cryptosporidium sp. ini dapat dilihat pada gambar berikut.
Gambar 2. Siklus hidup Cryptosporidium sp.9
Pustaka : https://ejournal.unri.ac.id/index.php/JIK/article/view/4268
SIKLUS HIDUP KHUSUS CRYPTOSPORIDIUM Ciri-ciri utama siklus hidup C. parvum atau C. hominis adalah bahwa siklus ini dimulai dengan menelan ookista yang sepenuhnya bersporulasi dan tahan lingkungan. Setelah pengeluaran di usus kecil bagian atas, sporozoit yang dilepaskan menembus lapisan lendir dan menempel pada enterosit di sekitarnya, menyebabkan mereka membentuk vakuola parasitoforous di sekitar parasit, yang kemudian berdiferensiasi menjadi trofozoit. Ciri yang tidak biasa dari vakuola ini adalah bahwa ia terletak di dalam membran plasma sel inang, tetapi di luar sitoplasma sel inang, dipisahkan dari yang terakhir oleh yang disebut organel pengumpan dan konsentrasi khusus elemen sitoskeletal sel inang. Pembelahan mitosis parasit pada saat ini menghasilkan tipe I meront dan produksi 6 atau 8 merozoit. Merozoit menyerupai sporozoit. Mereka melarikan diri dari vakuola parasitophorous dan menempel pada enterosit di sekitarnya, membentuk siklus infeksi aseksual yang diperkuat. Atau, infeksi merozoit dapat menyebabkan meront tipe II, dan produksi 4 merozoit. Seperti halnya merozoit yang berasal dari meront tipe I, merozoit tipe II melarikan diri untuk menginfeksi enterosit di sekitarnya, menghasilkan makrogamont (betina) atau microgamont (jantan). Enam belas atau lebih microgametes dari microgamont dilepaskan dan masing-masing dapat
membuahi
macrogamont
untuk
membentuk
zigot
diploid,
yang
berdiferensiasi menjadi ookista. Meiosis kemudian menghasilkan 4 sporozoit yang terbentuk. Ini merupakan siklus seksual, produk akhir yang merupakan ookista berdinding tipis yang bersporasi penuh (-20%) yang mengupas dalam host dan menghasilkan autoinfeksi, atau ookista berdinding tebal (-80%) yang diekskresikan ke dalam lingkungan.
Ookista Bentuk berdinding tebal berdinding penuh (berdiameter -5 μm) ini tahan
terhadap paparan lingkungan yang berkepanjangan di berbagai sumber air, dan juga tahan terhadap banyak agen desinfektan yang umum digunakan [13] , termasuk encer pemutih, yang dapat digunakan saat mengisolasi yang layak. ookista dari sampel tinja. Protein dinding cryptosporidium oocyst (COWPs)
berperan
dalam
resistensi
lingkungan terhadap ini
dan apicomplexans
lainnya [14] . Reseptor permukaan Oocyst juga berperan dalam memastikan bahwa tahap parasit ini dekat dengan jaringan target inang di usus kecil.
Excystation
In vitro , garam empedu dan suhu 37 ° C sangat efektif dalam menginduksi eksistasi,
sedangkan
enzim
pankreas
tidak. Apa
yang
disebut
spesies
lambung, Cryptosporidium muris (C. muris) dan Cryptosporidium andersoni (C. andersoni) , distimulasi untuk unggul secara in vitro di lingkungan asam atau di hadapan asam taurocholic, sedangkan spesies yang unggul di usus kecil bagian atas, C.
hominis dan C.
terakhir . Namun,
asam
parvum ,
hanya
taurocholic
unggul
sangat
dengan
efektif
jika
stimulus
yang
ookista
pra-
diasamkan. Enzim yang diturunkan dari sporozoit juga memainkan peran kunci dalam proses eksistasi.
Sporozoit
Sporozoit berbentuk spindel (-4 × 0,6 μm). Seperti halnya parasit apicomplexan lainnya, kompleks apikal sporozoit memainkan peran penting dalam motilitas meluncur yang digunakan oleh parasit untuk mengakses sel target, perlekatan sel target, dan pembentukan vakuola parasitoforous. Enzim sporozoit membantu lewat melalui selimut lendir. C. parvum sporozoit bergerak dengan gerakan meluncur yang ditenagai oleh motor aktin-myosin parasit, meninggalkan jejak terdiri dari komponen yang disekresikan oleh mikronem kompleks apikal, yang juga berkontribusi pada pemilihan sel inang. Studi molekuler dan proteomik telah mengidentifikasi beberapa protein sporozoit dengan peran potensial dalam motilitas, dan dalam adhesi sel dan invasi sel inang.
Adhesi pada sel inang dan pembentukan vakuola parasitophorous
Propulsi ke depan dari sporozoit menghasilkan pelekatan pada permukaan apikal enterosit. Ada beberapa kandidat protein parasit kompleks apikal (asal mikronem, rhoptry, dan granula padat) yang mungkin berperan dalam perlekatan. Protein
terkait membran sporozoit, CP47, adalah salah satu protein yang mengikat reseptor pada sel target seperti p57 glikoprotein yang terletak di perbatasan sikat ileum.Setelah lampiran telah terjadi, ada gerakan umum dari mikronem dan butiran padat dengan ekstensi rhoptry menuju situs lampiran. Banyak penelitian ultrastruktural menunjukkan bahwa sporozoit yang melekat dilalap oleh sel inang dan microvilli elongate yang berdekatan. Ini menelan parasit dan pembentukan vakuola parasitophorous melibatkan pembengkakan sel inang sebagai aquaporin I dan SGLTI symporter natrium-glukosa direkrut ke antarmuka host-parasit sel. Ada juga renovasi besar sitoskeleton di daerah yang melibatkan polimerisasi aktin sel inang. Jalur pensinyalan yang terlibat dalam proses invasi dan pembentukan vakuola parasitofor telah dipelajari secara luas. Sementara sebelumnya diyakini bahwa vakuola parasitophorous berasal dari sel inang, sekarang jelas bahwa ia memiliki kontribusi parasit yang signifikan. Selain pita padat elektron pada antarmuka sel vakuola sitoplasma-parasitoforous, parasit plasmalemma invaginate langsung di atas jaringan terminal, kompres, dan menjadi struktur membran yang sangat terlipat yang sering disebut sebagai organel pengumpan. Struktur ini diasumsikan untuk membentuk jalur utama untuk akses parasit ke nutrisi dalam sitoplasma sel inang, sebuah asumsi yang didukung oleh kehadiran protein pengikat kaset-ABC. Jalur langsung ke nutrisi sel inang sangat
diperlukan
untuk
parasit
ini. Genom
dari C.
parvum dan C.
hominis menunjukkan pemadatan dan kedua spesies tampaknya memiliki kemampuan biosintesis yang terbatas dan karena itu harus bergantung pada sel inang untuk nutrisi.
Trofozoit (diameter 1,5-2,5 μm)
Setelah penutupan sporozoit dalam vakuola parasitofor, dediferensiasi parasit mulai terjadi, diikuti oleh diferensiasi. Seperti halnya sporozoit yang menempel, terdapat gangguan dan pemanjangan microvilli sel inang yang berbatasan langsung dengan trofozoit.
Tipe I meront
Mitosis dalam trofozoit mengawali pembentukan meront tipe I di mana 8 atau 6 merozoit keluar dari tubuh residual yang terletak di dekat persimpangan sel inangparasit dan organel pengumpan. Seperti halnya trofozoit, mikrovili yang bersebelahan dengan tipe I meront rusak dan memanjang. Merozoit diproduksi oleh tunas dari tubuh residu schizont dan memanjang. Membran vakuol parasitophorous pecah dan merozoit tipe I lolos.
Tipe I merozoite
Tahap ini seperti batang (0,4 × 1,0 μm), dengan daerah apikal runcing. Motilitas, perlekatan pada membran apikal enterosit yang berdekatan dan pembentukan trofozoit (yaitu pembentukan siklus aseksual) umumnya dianggap sama atau sangat mirip dengan sporozoit. Sebagai contoh, CpSUB1, protein serine seperti sutilisin yang diduga berperan dalam invasi, ditemukan di kutub apikal sporozoit dan merozoit.
Tipe II meront
Sementara merozoit tipe I melanjutkan untuk menghasilkan lebih banyak tipe I meronts dalam serangkaian siklus aseksual, beberapa merozoit tipe I menghasilkan meront tipe II. Tahap ini memiliki ukuran mulai dari 3-5 μm. Perbedaan utama antara ini dan tipe I meront adalah bahwa hanya 4 merozoit yang
berkembang
pada
tahap
ini. Seperti
semua
tahap
intraseluler Cyptosporidium , ada organel pengumpan yang berkembang dengan baik.
Jenis merozoite II
Merozoit yang dilepaskan dari tipe II meronts kurang seragam dalam bentuk, sedikit lebih besar dan kurang aktif daripada yang dirilis dari tipe I meronts.
Microgamont dan macrogamont
Sementara beberapa merozoit tipe II memasuki enterocyte dan menghasilkan makrogamont, struktur bulat sampai oval berdiameter 4 hingga 6 μm dengan inti sentral yang besar, yang lain menghasilkan microgamont. Nukleus inti dari tubuh residu microgamont untuk membentuk 16 microgamet non-flagellated terpisah seperti batang (1,4 × 0,5 μm). Tahap bebas flagel ini keluar dari microgamont dan membuahi makrogamont yang berdekatan, menghasilkan satu-satunya tahap diploid dalam siklus hidup, zigot.
Zigot
Zigot berkembang menjadi ookista, kemudian mengalami sporogami, semuanya masih melekat pada membran apikal enterosit. Saat ookista berdiferensiasi, ia menjadi ookista berdinding tipis, atau ookista berdinding tebal. Mereka yang berkembang menjadi ookista berdinding tebal mengandung tipe I dan tipe II badan pembentuk dinding. Setelah mereka berdiferensiasi, ookista terlepas ke dalam lumen untuk menginfeksi inang dalam kasus ookista berdinding tipis, atau diekskresikan ke lingkungan dalam kotoran dalam kasus ookista berdinding tebal. Adalah mungkin tetapi tidak mungkin bahwa, di samping siklus kehidupan yang dijelaskan di atas, tahapan ekstraseluler gratis mungkin ada. Sementara mereka telah dijelaskan dalam model kultur sel in vitro, tidak ada deskripsi tahap ekstraseluler seperti itu dalam model in vivo .
Pustaka : https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3368497/