![Perbedaan Bakteri Dan Archaea PDF [PDF]](https://pdfs.asia/img/200x200/perbedaan-bakteri-dan-archaea-pdf.jpg)
6 0 2 MB
PR MIKROBIOLOGI “PERBEDAAN DINDING SEL BAKTERI DAN ARCHAEA”
Oleh: NAMA NIM MATA KULIAH DOSEN PROGRAM STUDI JENJANG
: : : :
SUSILAWATI. S ACD 214 004 MIKROBIOLOGI Dr. YANETRI ASI NION, M.Si Dr. LISWARA NENENG, M. Si : PENDIDIKAN BIOLOGI : MAGISTER PENDIDIKAN (S-2)
PROGRAM PASCASARJANA PENDIDIKAN BIOLOGI UNIVERSITAS PALANGKARAYA BULAN OKTOBER TAHUN 2015
1
1. BAKTERI a. Struktur sel Bakteri dan Bagian Bagian Sel Bakteri Struktur sel bakteri pada umumnya semuanya hampir sama yaitu nukleous yang berbentuk nukleoida. adanya dinding sel bakteri, serta kromosom minor berupa plasmid dan terdapat pilus atau pili dan ribosom. membran plasma, dan mesosom. Tambahan lain seperti flagellum (flagella), lapisan S,karboksisome, magnetosome dapat anda temukan pada bakteri tertentu. Berikut gambar struktur sel bakteri:
Struktur Sel Bakteri Dinding sel bakteri tersusun atas peptidoglikan. Peptidoglikan adalah rantai polisakarida yang tersusun atas Asam N-Asetilmuramik (NAM) dan NAsetilglukosamin (NAG). Dinding sel bakteri berfungsi dalam mempertahankan bentuk sel, menjaga tekanan dalam sel atau turgor sel dikarenakan banyaknya kandungan protein dalam sel. Dinding sel pada bakteri terbagi atas dua jenis yaitu gram positif dan gram negatif. Perbedaan keduanya terletak pada jumlah peptidoglikan dan karakteristik khusus pada membran sitoplasmanya. Membran Plasma Bakteri atau membran sitoplasma, Pada umumnya tersusun atas phospholipid bilayer. Akan tetapi khusus untuk bakteri gram negatif, memiliki membran terluar dan periplasma sebelum mencapai membran sitoplasma sehingga terlihat seperti tiga lapisan. Lapisan membran luar tersusun atas
2
lipopolisakarida dan protein. Hal ini yang membedakan antara gram negatif dan bakteri gram positif.
Struktur Dinding Sel bakteri
Lapisan membran sel bakteri Flagella atau flagellum (tunggal) pada sel bakteri berfungsi sebagai alat gerak bakteri. Terdapat beberapa macam flagella pada bakteri berdasarkan letaknya dan jumlahnya pada sel bakteri tersebut. Untuk lebih jelasnya tentang pembagian sel bakteri berdasarkan flagellanya silahkan baca pada artikel motilitas bakteri ini.
3
Fimbriae atau rambut sel merupakan tabung protein yang berada pada bagian luar sel. Fimbriae utamanya berfungsi dalam pelekatan akan tetapi pada beberapa kasus seperti Myxococcus dapat berfungsi dalam motilitas bakteri untuk membentuk badan buah. Selain fimbriae, ada juga pili, memiliki struktur yang sama dengan fimbriae akan tetapi lebih panjang dan lebih sedikit jumlahnya pada bakteri. Hal ini karena fungsinya yang lebih spesifik untuk konjugasi serta memperkuat pelekatan bakteri. Bagian dalam sel bakteri terdapat ribosom yang berperan dalam sintesis protein. Ribosom pada sel bakteri berbeda dengan sel eukariot dari segi ukuran. Ukuran ribosom bakteri adalah 70 S (S = Svedberg), dengan subunit besar 50 S dan subunit kecil 30 S. Selain ribosom juga terdapat sitoskeleton. Fungsi sitoskeleton pada sel bakteri sama dengan sitoskeleton pada eukariot. Terdapat juga inklusi pada sel bakteri seperti droplet lemak, kristal, glikogen, dan pigmen. Inklusi sel bakteri tidak memiliki membran dan tidak melakukan proses metabolisme. Selain itu, terdapat juga vakuola gas. Akan tetapi vakuola gas hanya terdapat pada bakteri tertentu seperti bakteri plankton dan cyanobakteria. Sel bakteri juga memiliki DNA yang berbentuk sirkuler dan pada beberapa bakteri terdapat DNA yang linear. Selain itu, bakteri juga memiliki plasmid atau DNA mini yang berfungsi dalam meneruskan informasi tertentu. Plasmid sering disebut sebagai ekstrakromosomal DNA. Mesosome yang sebelumnya dikenal sebagai sebuah pelipatan membran yang dihasilkan oleh sel bakteri sebagai organel atau interseluler membran ternyata ditemukan sebagai sebuah artifak yang disebabkan oleh proses dalam melakukan pengamatan menggunakan mikroskop elektron. Akan tetapi, terdapat beberapa macam interselular membran pada bakteri seperti Kromatofor pada bakteri fototrofik, yang mengandung bakterioklorofil dan karotenoid. Selain itu terdapat juga mikrokompartemen pada bakteri seperti karboksisome dan magnetosome. Mikrokompartemen berfungsi dalam metabolisme tertentu pada bakteri tertentu dan dibungkus oleh protein sehingga tidak mengandung lipid karena tidak dibungkus oleh membran. Sepertinya hampir semua bagian dan struktur bakteri sudah anda pelajari. Untuk lebih jelasnya saya sarankan anda baca
4
buku khusus tentang bakteri seperti buku bakteriologi. Bakteri merupakan organisme prokariotik
yang
memiliki dinding
sel yang tersusun dari
peptidoglikan. Hal ini berbeda dengan tumbuhan yang dinding selnya tersusun dari selulosa, pektin, maupun lignin. Dinding sel bakteri memiliki struktur yang agak kaku yang terletak di luar membran sel. Peranan dinding sel tersebut adalah untuk mempertahankan bentuk sel dan mencegah sel mengalami lisis. Komponen utama dari dinding sel bakteri adlah peptidoglikan atau disebut juga dengan murein. Peptidoglikan merupakan suatu polimer yang berukuran besar yang dihubungkan dengan ikatan kovalen. Peptidoglikan terdiri dari dua macam derivat polisakarida, yakni N-acetylglucosamine dan N-acetylmuramic acid serta asam amino seperti L-alanine, D-alanine, dan D-glutamic acid. Pada polimer peptidoglikan molekul N-acetylglucosamine bergantian dengan molekul N-acetylmuramic acid yang saling berpaut silang membentuk glycan tetrapeptide (Gambar 1). glycan tetrapeptide ini melalui tetrapeptida, empat asam amino. Sebagian besar bakteri gram positif memiliki asam amino ketiga berupa lisin sedangkan sebagian besar bakteri negatif berupa asam diaminophimelat.
Gambar 1. Struktur peptidoglikan yang setiap unit mengalami repetisi membentuk glycan tetrapeptide. Rantai panjang dari peptidoglikan ini disintesis secara berdampingan untuk membentuk suatu lembaran yang melingkupi suatu sel (Gambar paling
5
atas). Rantai-rantai peptidoglikan tersebut dihubungkan secara menyilang oleh asam amino. Sementara ikatan glikosida menghubungkan gula di dalam untaian glikan secara kovalen (Gambar 2).
Gambar 2. Struktur peptidoglikan pada Escherichia coli dan Staphylococcus aureus. Pada dinding sel bakteri gram positif memiliki molekul tambahan berupa asam teikoat yang terdiri atas gliserol, fosfat, dan ribitol gula alkohol dalam bentuk polimer dengan panjang 30 unit. Polimer-polimer tersebut terkadang memanjang sampai keluar dari dinding sel dan kapsul (Gambar 3). Pada bakteri gram positif memiliki lapisan peptidoglikan yang relatif tebal dengan ukuran 2080 nm. Lapisan peptidoglikan tersebut menempel pada permukaan luar membran sel. Bakteri jenis ini tidak memiliki membran luar maupun ruang periplasmik. Sehingga dengan menggunakan pewarnaan gram (Hans Christian Gram), maka bakteri ini akan nampak berwarna ungu.
6
Gambar 3. (a) struktut asam teikoat; (b) ilustrasi dinding sel bakteri gram positif. Adapun pada bakteri gram negatif memiliki struktur yang lebih kompleks dibandingkan dengan bakteri gram positif. Komposisi peptidoglikan sekitar 1020% dan sisanya berupa polisakarida, protein, dan lipid. Dinding sel terdiri atas membran luar yang menyusun permukaan luar dinding dan berbatasan dengan ruang periplasmik yang sangat sempit (Gambar 4). Pada pewarnaan gram, bakteri ini tidak bisa mempertahankan warna kristal violet pada tahap dekolorisasi. Hal ini dikarenakan dinding selnya sangat tipis dan jumlah lipoprotein serta lipopolisakarida banyak pada dinding sel.
Gambar 4. Diagram dinding sel bakteri gram negatif.
7
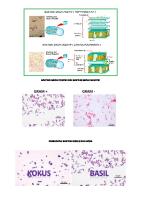
Perbedaan antara bakteri gram positif dan negatif disajikan dalam Gambar 5 dan Tabel 1 berikut ini.
Gambar 5. Perbedaan gram positif dan gram negatif. Tabel 1. Perbedaan bakteri gram (+) dan gram (-). Karakteristik Bakteri Gram Bakteri Gram (-) (+) Peptidoglikan Lapisan tebal Lapisan tipis Asam Teikoat Sering dijumpai Tidak ada Membran Luar Tidak ada Ada Ruang Periplasmik Tidak ada Ada Bentuk Sel Kaku Kaku dan fleksibel Hasil Pencernaan Enzim Protoplas Spheroplast Sensitivitas terhadap Paling sensitif Kurang sensitif Warna dan Antibiotik 2. ARCHAEA 2.1. Bentuk dan Ukuran Sel Secara umum struktur sel Archaea memiliki bentuk yang hampir sama seperti bakteri, dan bentuknya cukup beragam. Beberapa Archaea berbentuk batang/basil, bulat/kokus, atau spiral. Bahkan terdapat beberapa Archaea yang memiliki bentuk “tidak biasa” , yaitu segitiga dan persegi panjang. Meskipun morfologi sel relatif mudah untuk diamati, tetapi terkadang sulit untuk
8
membedakan bakteri dan Archaea, karena keduanya memiliki ragam bentuk yang hampir sama.
Gambar 3. Beberapa bentuk morfologi yang terdapat pada Archaea (a) Methanobrevibacter smithii; (b) Methanobacterium uliginosum; (c) Methanosphaera stadtmanae; (d) Methanoplanus limicola ; (e) Methanospirillum hungatei; (f) Halobacteriumhalobium; (g) Halococcus morrhuae; (h) Thermoplasma acidophilum; (i) Methanolobus vulcani; (j) Pyrococcus furiosus; (k) Haloferax mediterranei; (l) Thermofilum ‘librum’; (m) Pyrodictium occultum; (n) Thermoproteus tenax. Archaea merupakan organisme yang berukuran sangat kecil, yaitu sekitar 1.5-2.5 µm (Beveridge, 2001). Ukuran yang kecil ini memberikan keuntungan tersendiri bagi sel tersebut. Sel yang berukuran lebih kecil memiliki luas permukaan yang lebih besar dibandingkan dengan volume sel, jika dibandingkan dengan sel yang berukuran lebih besar. Sehingga memiliki rasio permukaan terhadap volume lebih tinggi. Rasio permukaan/volum memberikan beberapa akibat pada kehidupannya. Sebagai contoh pada pertukaran nutrisi, sel yang memiliki rasio permukaan/volum lebih tinggi akan mendukung pertukaran nutrisi lebih cepat dibanding yang lebih rendah, oleh karena itu sel yang lebih kecil akan tumbuh lebih cepat dibandingkan dengan sel yang lebih besar karena memiliki rasio yang lebih tinggi. Sedangkan secara genetik, hal ini dapat berdampak pada evolusi karena sel Archaea adalah haploid, sehingga mutasi akan diekspresikan
9
secara langsung. Sedangkan mutasi itu sendiri adalah sumber dari suatu evolusi. Oleh sebab itu Archaea dapat lebih cepat menanggapi perubahan lingkungan.
2.2 Membran sitoplasma pada Archaea Struktur dasar dari membran sel Archaea tersusun atas fosfolipid. Struktur ini tersusun dari molekul gliserol yang berikatan dengan fosfat pada ujung pertama (kepala) dan berikatan dengan rantai samping yang berupa isoprenoid pada ujung lainnya (ekor).
Gambar 4. membran sel Archaea. Karena sifatnya yang hidrofilik maka ketika membran sel berada pada lingkungan cair, ujung molekul yang mengandung gugus fosfat akan berada pada permukaan luar membran yang berhubungan langsung dengan lingkungan luar sel, dan sisi lainnya yang bersifat hidrofobik akan berada dibagian dalam. Pelapisan seperti ini menciptakan penghalang kimia yang sangat efektif disekitar sel dan membantu dalam menciptakan keseimbangan kimiawi. Secara komposisi, membran sel Archaea memiliki perbedaan dengan membran sel bakteri dan eukaria. Perbedaan tersebut antara lain adalah perbedaan kiralitas gliserol yang menjadi penyusun membran sel, ikatan antara gliserol dan rantai samping isoprenoid berupa ikatan eter, rantai samping berupa isoprenoid bukan asam lemak seperti pada bakteri dan eukaria, dan memiliki rantai samping yang bercabang.
10
2.2.1. Kiralitas dari gliserol Gliserol yang digunakan Archaea untuk membentuk fosfolipid merupakan stereoisomer dari gliserol yang digunakan untuk membentuk membran sel pada bakteri dan eukaria. Dua molekul yang stereoisomer adalah cerminan satu sama lain. Pada membran sel bakteri dan eukaria, gliserol yang menyusun membran selnya berupa D-Gliserol, sedangkan pada arkaea berupa L-gliserol.
Gambar 5. Struktur penyusun membran sel Archaea dan bakteri/eukaria. 2.2.2. Ikatan eter Pada kebanyakan organisme, gliserol yang terdapat pada membran selnya akan berikatan dengan rantai samping menggunakan ikatan ester. Namun tidak demikian halnya pada membran sel Archaea. Ikatan yang terbentuk antara gliserol dan rantai samping pada membran sel Archaea adalah ikatan eter. Hal ini memberikan fosfolipid yang dihasilkan memiliki sifat mekanik kimia yang berbeda dari lipid membran organisme lain.
Gambar 6. Ikatan yang terbentuk pada membran sel bakteri/eukaria dan Archaea.
11
2.2.3. Rantai Isoprenoid Archaea memiliki rantai samping penyusun fosfolipid yang berbeda dengan bakteri dan eukaria. Rantai samping penyusun fosfolipid pada bakteri dan eukaria adalah asam lemak, sedangkan pada Archaea rantai samping yang dimilikinya adalah isoprenoid. Isoprenoid merupakan hidrokarbon yang memiliki 20 atom C dan merupakan anggota paling sederhana dari kelas bahan kimia yang disebut terpene. Menurut definisi, terpene adalah molekul yang menghubungkan molekul isoprenoid bersama-sama.
Gambar 7. Struktur membran monolayer pada Archaea. Lipid yang terdapat pada Archaea termoasidodfil dan metanogen adalah tetralipid, dimana ujung rantai samping phytanil pada struktur tetralipid berikatan secara kovalen dengan molekul gliserol yang lain. Sehingga akan membentuk struktur monolayer. Struktur seperti ini tidak memiliki area tengah yang kosong seperti pada struktur lipid bilayer. Sehingga struktur seperti ini memiliki resistensi yang lebih terhadap temperatur tinggi dibandingkan struktur lipid bilayer. Pada umumnya Archaea yang hidup optimal pada suhu tinggi, membran selnya terdiri dari lipid monolayer ataupun kombinasi antara lipid bilayer dan monolayer.
Gambar 8. Membran monolayer dan bilayer.
12
2.2.4. Rantai samping yang Bercabang Tidak hanya rantai samping Archaea yang dibentuk dari komponen yang berbeda. Akan tetapi rantai sampingnya memiliki struktur fisik yang juga berbeda. Rantai samping pada membran sel Archaea memiliki cabang, karena penggunaan isoprenoid untuk membentuk rantai sampingnya. Asam lemak pada bakteri dan eukariot tidak memiliki rantai cabang, sehingga sifat ini menjadikan membran Archaea yang memiliki karakter unik. Hal ini menciptakan beberapa properti yang menarik di membran Archaea. Rantai samping isoprenoid bisa bergabung bersama-sama antara dua rantai samping fosfolipid tunggal atau bergabung ke rantai fosfolipid sisi lain di sisi lain membran (membentuk fosfolipid transmembran). Rantai samping tersebut juga dapat mempunyai kemampuan untuk membentuk cincin karbon. Hal ini terjadi ketika salah satu cabang mengelilingi dan mengikat atom bawah rantai untuk membuat cincin lima atom karbon. Cincin tersebut diperkirakan memberikan stabilitas struktural membran.
2.3 Dinding Sel Archaea Archaea memiliki keragaman dalam hal lapisan yang menyelubungi selnya. Beberapa Archaea memiliki lapisan protein permukaan atau S-layer. Lapisan ini terdiri dari protein monomolekular yang identik atau lebih dikenal dengan sebutan glikoprotein (Kandler dan Konig, 1993). Lapisan ini secara langsung berhubungan dengan bagian luar membran plasma dan berfungsi untuk melindungi dari lisis osmotik. Lapisan ini juga dapat berfungsi sebagai penyeleksi molekul yang dapat masuk kedalam sel.
Gambar 9. S-Layer.
13
Selain S-Layer, diketahui beberapa Archaea juga memiliki struktur yang mirip dengan dinding sel pada bakteri, namun berbeda dalam hal komposisi kimia penyusunnya. Dinding sel Archaea tidak memiliki peptidoglikan namun memiliki molekul yang mirip dengan peptidoglikan yang disebut pseudomurein. Pseudomurein dibangun dari N-Asetil glukosamin dengan Asam N-Asetil talosamin uronat yang berikatan dengan ikatan glikosidik pada β-1,3 hal ini berbeda dengan peptidoglikan pada bakteri yang dibangun menggunakan N-Asetil glukosamin dan N-Asetil muramat yang berikatan pada β-1,4.
Gambar 10. Struktur pseudomurein. Perbedaan lainnya adalah asam amino yang terdapat pada pseudomurein semuanya berupa L-Steroisomer. Struktur seperti ini memberikan dampak yang menguntungkan pada Archaea, yaitu dinding sel mereka resisten terhadap antibiotik dan juga tidak terpengaruh terhadap aktivitas lisosim dan protease yang umum (Konig, 2001). Beberapa Archaea tidak memiliki pseudomurein namun memiliki
polisakarida
lainnya,
yaitu
glutaminylglycan,
heterosakarida,
methanochondroitin.
2.4. Struktur Permukaan Sel Archaea, Inklusi Sel, dan Vesikula Udara Penelitian mengenai struktur tambahan pada permukaan sel Archaea telah banyak dilakukan dengan memanfaatkan observasi elektron mikroskopis pada beberapa jenis Archaea. Penelitian ini menunjukkan beberapa tipe struktur tambahan pada permukaan sel Archaea, seperti pili dan flagella yang tampak
14
seperti struktur yang ada pada bakteri, tetapi ternyata memiliki perbedaan. Selain itu struktur lain seperti cannulae (kanula), Hami, Iho670 Fibers, dan bindosome muncul
sebagai
struktur
unik
lain
yang
dimiliki
oleh
Archaea.
2.4.1. Struktur Permukaan Sel Archaea a. Pili Fimbriae dan pili merupakan struktur filamen yang tersusun atas protein yang memanjang dari permukaan sel dan memiliki banyak fungsi. Fimbriae memungkinkan sel untuk menempel pada suatu permukaan. Secara umum pili mirip dengan fimbriae, tetapi pili lebih panjang dan hanya satu atau sebagian kecil pili yang bisa melekat pada permukaan sel. Fungsi pili itu sendiri adalah untuk memfasilitasi pertukaran gen di antara sel pada suatu proses yang disebut sebagai konjugasi. Walaupun sebenarnya proses konjugasi tidak selalu diperantarai oleh pili.
Gambar 11. Tanda panah menunjukkan pili pada struktur permukaan sel. b. Cannula, Hami, Iho670 Fibers, dan Bindosome Struktur permukaan sel Archaea terdiri dari banyak bagian, yaitu kanula, hami, Cannula, Hami, Iho670 Fibers, dan Bindosome. Struktur permukaan tersebut tidak banyak dibahas seperti halnya pili dan flagella, hal ini disebabkan karena sistem genetik di dalam struktur tersebut tidak mudah untuk dipelajari dan tidak ditemukan pada semua jenis Archaea. c. Cannulae (Kanula) Kanula merupakan jaringan tubula yang sampai saat ini hanya ditemukan pada genus Pyrodictium. Kanula berupa pipa berongga berdiameter luar 25 nm (Gambar 12) yang sangat resisten terhadap panas dan proses denaturasi (Rieger et
15
al., 1995). Strukturnya hampir sama dengan struktur permukaan sel lainnya yaitu terbentuk atas lapisan glikoprotein, yang memiliki tiga subunit glikoprotein yang homolog. Kanula menunjukkan aktivitasnya sebagai penghubung intraseluler antar ruang periplasmik sel yang berbeda (Nickell et al., 2003). Walaupun fungsi kanula belum diketahui secara jelas, tetapi dapat diasumsikan bahwa dengan adanya kanula, sel dapat melakukan pertukaran nutrisi atau bahkan materi genetik.
Gambar 12. Kanula (Rieger et al., 1995). d. Hami Struktur permukaan Archaea yang lain adalah hamus atau hami (Gambar 13). Hami banyak ditemukan pada Archaea yang hidup di daerah suhu rendah yang mengandung
kadar
sulfat
tinggi
(cold
sulphidic
springs).
Strukturnya
menunjukkan filamen-filamen yang sangat kompleks dengan kenampakan seperti kawat berduri yang ujungnya memiliki kait dengan diameter 60 nm (Moissl et al., 2005). Masing-masing sel dikelilingi oleh sekitar 100 hami. Hami stabil pada kisaran temperatur dan pH yang luas yaitu antara 0-70 oC dan 0,5-11,5. Hami dapat bertindak sebagai perantara proses adesi seluler permukaan terhadap komposisi kimia yang berbeda sebagaimana adesi yang berlangsung di antara sel. Hami juga terbukti menjadi komponen protein utama dalam pembentukan biofilm Archaea, dimana sel membentuk susunan tiga dimensi yang jaraknya konstan melalui proses perlekatan antar sel Archaea (Henneberger et al., 2006).
16
Gambar 13. (a) Sekitar 100 hami keluar secara melingkar di permukaan sel. (b) Kenampakan kait yang berada di ujung hami. Tanda panah menunjukkan lokasi kait. (c) Hami menunjukkan kenampakan seperti kawat berduri (Moissl et al., 2005). e. Bindosome Bindosome (Gambar 14) adalah struktur Archaea yang diduga mempunyai fungsi unik pada Sulfolobus solfataricus (Albers dan Pohlschröder, 2009). Komponen struktural bindosome yang utama adalah substrat pengikat protein (substrat binding protein/SBP) yang diketahui sebagai glikoprotein (Elferink et al., 2001), yang disusun oleh Pilin tipe IV seperti pada sekuen peptida sinyal dan mengandung protein khas yang diketahui mampu membentuk struktur oligomerik pada Archaea dan bakteri. Susunan oligomerik komplek berperan dalam penyerapan gula, hal ini dapat membantu S. solfataricus untuk dapat tumbuh pada substrat yang bervariasi (Ng et al., 2008).
Gambar 12. Gambar asli bindosome belum diketahui secara pasti, dan gambar diatas merupakan formasi alternatif yang menunjukkan bindosome terletak pada S-layer (Ng et al., 2008).
17
f. Iho670 Fibers Pada pertengahan tahun 2009 telah dilakukan penelitian oleh Muller et al. mengenai struktur permukaan Ignicoccus hospitalis, hasilnya menunjukkan adanya tambahan permukaan sel baru yang kemudian diberi nama Iho670 fiber (Gambar 15). Iho670 fiber merupakan struktur yang sangat rapuh, berbeda dengan flagella dan pili yang memliliki struktur primer dari protein. Hal ini juga menunjukkan bahwa Iho670 fiber bukan salah satu organel sel yang motil. yang menjadi bagian menarik adalah bahwa komponen utama Iho670 fiber disintesis oleh Pilin tipe IV seperti peptida sinyal dan diproses oleh peptidase prepilin homolog. Karena Pilin tipe IV seperti sistem ini juga digunakan untuk flagela, pili tertentu, dan bindosome dalam Archaea, Pilin tipe IV menjadi jalur yang sangat banyak digunakan oleh Archaea dalam hal perakitan struktur permukaan.
Gambar 13. Hasil analisis serat Ignicoccus hospitalis menggunakan TEM (Transmission Electron Microscopy) yang mengindikasikan adanya Iho670 fibers. 2.4.2. Inklusi Sel Di dalam sel prokariotik biasanya terdapat senyawa lain yang menyertai sel di dalam sitoplasma yang disebut dengan inklusi sel. Inklusi sel berfungsi sebagai energi cadangan atau sebagai tempat penyimpanan struktur building blocks. Penyimpanan karbon atau senyawa lain di dalam inklusi yang tidak larut dalam air bermanfaat bagi sel karena dapat mengurangi tekanan osmotik yang dapat mungkin terjadi apabila senyawa dalam jumlah yang sama terlarut dalam sitoplasma (Madigan et al., 2012).
18
Gambar 16. Tanda panah menunjukkan poly-β-hydroxyalkanoat (PHA) (Madigan et al., 2012). Salah satu jenis inklusi sel yang paling banyak ditemukan di dalam organ prokariotik adalah asam poly-β-hydroxybutirat (PHB). PHB adalah lipid yang tersusun atas unit-unit asam β-hydroxybutirat. Sedangkan polimer yang diproduksi oleh Archaea adalah poly-β-hydroxyalkanoat (PHA) (Gambar 16). PHA disintesis oleh Archaea di dalam polimer penyimpanan ketika sel mengalami kondisi pertumbuhan yang tidak seimbang. PHA merupakan salah satu jenis komoditas plastik yang dapat dirombak menjadi karbondioksida dan air melalui proses mineralisasi mikrobiologis secara alami.
2.4.3. Vesikula Udara Salah satu jenis Archaea yang bersifat planktonic dan mampu hidup di air laut adalah Nitrosopumilus maritimus dari kelompok Crenarchaeota (BrochierArmanet et al., 2011). Jenis organisme ini mampu mengapung di air laut karena memiliki
vesikula
udara.
Kemampuan
mengapung
yang
dimilikinya
memungkinkan untuk menempatkan diri dalam kolom air untuk dapat merespon kondisi lingkungan.
19
Gambar 17. Vesikula udara pada struktur permukaan sal Archaea. Secara umum struktur vesikula udara tersusun atas protein yang berbentuk kumparan, berongga namun kaku dengan panjang dan diameter yang bervariasi (Gambar 17). Panjang vesikula udara yang dihasilkan oleh masing-masing organisme berbeda-beda, mulai dari 300 sampai lebih dari 1000 nm dengan lebar 45 sampai 120 nm, tetapi kisaran ukuran tersebut masih bisa berubah-ubah. Jumlah vesikula dalam satu organisme sangat bervariasi mulai dari sedikit hingga ratusan tiap selnya, kedap air dan larut dalam gas (Madigan et al., 2012).
2.5 Pergerakan Sel Archaea a. Flagella Archaea Flagella Archaea berukuran sangat kecil hingga mencapai setengah dari ukuran flagella bakteri, yaitu 10-13 nm (Madigan et al., 2012). Flagella Archaea memberikan kemampuan terhadap sel Archaea untuk dapat bergerak memutar seperti halnya bakteri. Flagella Archaea tidak hanya sebagai alat untuk bergerak, tetapi juga berperan dalam interaksi di dalam sel dan sebagai pengenal pada permukaan sel sebagai syarat terbentuknya biofilm pada beberapa Archaea. Flagella ditemukan pada semua sub kelompok utama Archaea Crenarchaeota dan Euryarchaeota yaitu halofil, haloalkalofil, metanogen, hipermetrofil, dan termoasidofil. Sampai saat ini telah dilaporkan berbagai macam Archaea yang memiliki
flagella,
termasuk
Methanococcus,
Halobacterium,
Natrialba, Thermococcus dan Pyrococcus (Ng et al., 2006).
20
Sulfolobus,
Gambar 18. (a) Sel Methanococcus maripaludis dengan diameter 1μm menunjukkan banyaknya flagella yang terdapat di permukaan selnya dan (b) flagella yang telah dimurnikan dari sel Methanococcus maripaludis. Tanda panah menunjukkan kait di ujung flagella. Secara umum penampakan flagella Archaea (Gambar 18) mirip dengan flagella bakteri tetapi flagella Archaea memiliki pergerakan yang unik seperti pada pili bakteri tipe IV (Jarrell et al., 2007). Kemiripan ini meliputi struktur flagella termasuk keberadaan jumlah gen pada masing-masing struktur. Pada awal penelitian mengenai flagella Archaea, diketahui kemiripan antara flagella Archaea dengan pili bakteri tipe IV adalah pada N-termini (Faguy et al., 1994) dan adanya pilin tipe IV yang mirip sinyal peptide (Kalmokoff and Jarrell, 1991). Penelitian terbaru menyebutkan bahwa protein yang ada pada flagella Archaea maupun pili bakteri tipe IV adalah ATPase (Bayley and Jarrel, 1998), membran protein (Peabody et al., 2003) dan sinyal peptidase (FlaK/PibD) (Bardy and Jarrel, 2002). Salah satu perbedaan antara flagella Archaea dengan flagella bakteri diketahui pada penelitian yang dilakukan tahun 2008 oleh Streif et al. mengenai pergerakan memutar pada flagella Archaea, hasilnya menunjukkan bahwa pergerakan flagella tersebut didukung oleh proses hidrolisis ATP dan bukan dari proton
atau
natrium
seperti
yang
digunakan
oleh
flagella
bakteri.
b. Kemotaksis Archaea Kemotaksis merupakan respon gerakan Archaea terhadap rangsangan dari senyawa kimia. Walaupun Archaea termasuk ke dalam kelompok yang berbeda dari bakteri, tetapi banyak spesies Archaea yang memiliki sifat kemotaksis. Berbagai macam protein yang mengatur proses kemotaksis pada bakteri juga ditemukan pada Archaea yang mampu bergerak (motil).
21
2.6. Pengemasan DNA Archaea Dalam filogenetik Archaea berbeda dengan bakteri, walaupun keduanya memiliki beberapa kemiripan dalam struktur sel. Perbedaan ini lebih pada taraf molekular antara keduanya, dimana Archaea memiliki banyak kesamaan dengan eukaria. Salah satu contohnya adalah pengemasan DNA pada Archaea. DNA pada Archaea dikemas dalam bentuk sirkular, dimana pada beberapa Archaea pengemasannya melibatkan DNA-girase dan protein histon untuk membentuk struktur DNA superkoil. Hal ini berbeda dengan bakteri yang membentuk struktur DNA
superkoil
dengan
bantuan
DNA-girase
saja.
Pengemasan
DNA
menggunakan protein histon seperti ini mirip dengan pengemasan DNA pada eukaria. Protein histon yang ditemukan pada Archaea berukuran lebih pendek dibandingkan dengan protein histon eukaria, tetapi keduanya memiliki sekuen asam amino dan struktur 3 dimensi yang homolog. Pada beberapa Archaea juga ditemukan beberapa titik awal replikasi, dimana protein yang mengenali titik awal replikasi dan sintesis DNA memiliki banyak kemiripan dengan eukaria dibandingkan dengan bakteri. Selain itu Archaea juga memiliki beberapa RNA polimerase. Hal ini berbeda dengan bakteri yang hanya memiliki satu RNA polimerase. Faktor transkripsi yang dimiliki Archaea juga memiliki kemiripan dengan faktor transkripsi pada eukaria. Beberapa gen penyandi tRNA dan rRNA Archaea memiliki intron. Intron yang terdapat pada Archaea diproses dengan mekanisme yang sedikit berbeda dengan intron pada eukaria. Sedangkan pada bakteri tidak ditemukan intron. Pada saat sintesis protein Archaea membutuhkan ribosom yang fungsional serta beberapa faktor translasi. Ribosom yang terdapat pada Archaea mirip dengan ribosom pada bakteri, yaitu sama-sama 70S. Namun faktor translasi yang ditemukan pada Archaea ternyata dua kali lebih banyak dibanding dengan yang ada pada bakteri. Bakteri dan Archaea menggunakan asam amino yang berbeda pada awal proses translasi. Asam amino yang digunakan bakteri adalah N-formil metionin, sedangkan Archaea adalah metionin. Metionin juga merupakan asam amino yang digunakan eukaria untuk awal proses translasi. Secara keseluruhan,
22
perbandingan sekuen menunjukkan beberapa kesamaan antara eukaria dan Archaea dalam hal RNA dan protein yang digunakan untuk membentuk translation machine.
23
DAFTAR PUSTAKA Albers S dan Pohlschr¨oder M. 2009. Diversity of Archaeal type IV pilin-like structures,” Extremophiles, vol. 13, no. 3, pp. 403–410. Bardy SL dan Jarrell KF. 2002. Flak of the archaeon Methanococcus maripaludis possesses preflagellin peptidase activity, FEMS Microbiology Letters, vol. 208, no. 1, pp. 53– 59. Bayley DP dan Jarrell KF. 1998. Further evidence to suggest that Archaeal flagella are related to bacterial type IV pili,Journal of Molecular Evolution, vol. 46, no. 3, pp. 370–373. Brochier-Armanet, CP. Deschamps, P. López-García, Y. Zivanovic, F. RodríguezValera and D. Moreira. 2011. Complete-fosmid and fosmid-end sequences reveal frequent horizontal gene transfers in marine uncultured planktonic Archaea. The ISME Journal. Vol. 5. p.1291-1302. Elferink MGL, Albers S, Konings W, and Driessen AJM. 2001. Sugar transport in Sulfolobus solfataricus ismediated by two families of binding protein-dependent ABC transporters. Molecular Microbiology, vol. 39, no. 6, pp.1494–1503. Faguy DM, Jarrell KF, Kuzio J, and Kalmokoff ML. 1994. Molecular analysis of Archaeal flagellins: similarity to the type IV pilin—transport superfamily widespread in bacteria. Canadian Journal of Microbiology, vol. 40, no. 1, pp. 67– 71. Henneberger R, Moissl C, Amann T, Rudolph C, dan Huber R. 2006. New insights into the lifestyle of the cold-loving SM1 euryarchaeon: natural growth as a monospecies biofilm in the subsurface, Applied and Environmental Microbiology, vol. 72, no. 1, pp. 192–199. Jarrell KF, S. Y. Ng, and Chaban B. 2007. Flagellation and chemotaxis, in Archaea: Molecular and Cellular Biology, R. Cavicchioli, Ed., pp. 385–410, ASM Press, Washington, DC, USA. Kalmokoff ML dan Jarrell KF. 1991. Cloning and sequencing of a multigene family encoding the flagellins of Methanococcus voltae, Journal of Bacteriology, vol. 173, no. 22, pp. 7113–7125
24
Konig H. 2001. Archaeal cell wall. Di dalam: Encyclopedia of life science. Chichester : 1486-1493 Moissl C, Rachel R, Briegel A , Engelhardt H, and Huber R. 2005. The unique structure of Archaeal ’hami’, highly complex cell appendages with nanograppling hooks, Molecular Microbiology, vol. 56, no. 2, pp. 361–370. Muller, D.W., C. Meyer, S. Gurster, U. Kuper, H. Huber, R. Rachel, G. Wanner, R. Wirth, and A. Belack. 2009. The Iho670 fibers of Ignicoccus hospitalis: a new type of Archaeal cell surface appendage. Journal of Bacteriology. Vol. 191, No. 20. p. 6465–6468 Ng S.Y., B. Zolghadr, A.J.M. Driessen, S. J. Albers, and K. F. Jarelli. 2008. Cell surface structures of Archaea. Journal of Bacteriology. Vol. 190. No. 18. P. 6039–6047. Ng, S. Y., B. Chaban, and K. F. Jarrell. 2006. Archaeal flagella, bacterial flagella and type IV pili: a comparison of genes and posttranslational modifications. J. Mol. Microbiol. Biotechnol. 11:167–191. Nickell R, Hegerl R, Baumeister W, and Rachel R. 2003. Pyrodictium cannulae enter the periplasmic space but do not enter the cytoplasm, as revealed by cryoelectron tomography, Journal of Structural Biology, vol. 141, no. 1, pp. 34–42. Peabody CR, Chung YJ, Yen MR, Vidal-Ingigliardi D, Pugsley AP, and Saier MH. 2003. Type II protein secretion and its relationship to bacterial type IV pili and Archaeal flagella, Microbiology, vol. 149, no. 11, pp. 3051–3072. Rieger G,Rachel R, Hermann R, dan Stetter KO. 1995. Ultrastructure of the hyperthermophilic archaeon Pyrodictiumabyssi, Journal of Structural Biology, vol. 115, no. 1, pp. 78– 87. Streif S, Staudinger WF, Marwan W, and Oesterhelt D. 2008. Flagellar rotation in the archaeon Halobacterium salinarum depends on ATP, Journal of Molecular Biology, vol. 384, no. 1, pp. 1–8. Thoma C, Frank M, Rachel R. 2008. The Mth60 fimbriae of Methanothermobacter thermoautotrophicus are functional adhesins, Environmental Microbiology, vol. 10, no. 10, pp. 2785–2795.
25
Woese C, Kandler O, dan Wheelis ML. 1990. Towards a natural system of organisms: Proposal for the domains Archaea, Bacteria, and Eucarya. Proc. Nati. Acad. Sci. 8 Yuwono T. 2005. Biologi molekular. Safitria, editor. Jakarta : Erlangga.
26